

OSTALI SEMINARSKI RADOVI
IZ BIOLOGIJE : |
|||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||

Taksonomski kriteriji u sistematici virusa
Broj virusa kao patogena ili „tihih putnika“ drugih organizama, od bakterija do sisara, vrlo je velik. Otkrivanjem novih niša za život, te poboljšanjem osjetljivosti i specifičnosti raznih tehnika istraživanja, broj virusa se širi. Zbog specifičnih svojstava virusa (građe i aktivnosti), kriteriji u klasifikaciji koji vrijede za biljni i životinjski svijet, ne mogu se primjeniti u taksonomiji virusa.
Taksonomija virusa se stalno mijenja, počev od trivijalnih promjena (korištenjem kurzive za pisanje imena vrsta), pa sve do kompletne reorganizacije potaknute eksplozijom niza novih informacija. U težnji da se stvori jedinstvena klasifikacija virusa, 1973. godine osnovan je Međunarodni komitet za taksonomiju virusa (International Committee on Taxonomy of Viruses - ICTV). Danas, ICTV djeluje pod pokroviteljstvom Odjela za virologiju Međunarodne unije mikrobioloških društava. Međunarodni komitet za taksonomiju virusa je odbor koji odobrava i organizira taksonomsku klasifikaciju virusa. Oni su razvili univerzalnu taksonomsku shemu za viruse s ciljem da se opišu svi virusi živih organizama. Univerzalni sistem taksonomije virusa zasniva se na Linneovoj klasifikaciji na nivou reda, familije, podfamilije, roda i vrste.
U aktuelnom 8. Izvješću (2009.) ICTV klasifikacije i taksonomije, ustanovljeno je 6 redova (Caudovirales, Herpesvirales, Mononegavirales, Nidovirales, Picornavirales i Tymovirales). Dakle, u taksonomiji virusa trenutno postoji 6 redova, 86 familija, 19 podfamilija, 348 rodova, 2290 vrsta i oko 4,000 tipova koji su još neodređeni. S ciljem da se omogući lakše praćenje istraživanja sistematskih i evolucijskih odnosa između virusa, 1991. godine Odbor osniva i vlastitu bazu podataka – ICTVdB. Vremenom je ona postala glavni referentni izvor i istraživački alat za taksonomiju virusa, te sadrži taksonomske podatke za preko 2,000 vrsta virusa.
2. Definicija virusa
Virusi spadaju u grupu mikroorganizama acelularne građe. Nalaze se na najnižem stepenu organizacije živih sistema. To su živa bića koja su prostija od ćelija, zbog čega se često nazivaju i virusne čestice. Obzirom da sadrže nukleinsku kiselinu i proteine, virusi se ubrajaju u živa bića. Pošto se sami ne mogu razmnožavati, oni koriste žive ćelije i njihove metaboličke mehanizme za svoj rast i razmnožavanje. Ta sposobnost da se razmnožavaju u živoj ćeliji i stvaraju sebi slično potomstvo čini ih živim bićima, a korištenje ćelije domaćina čini ih striktnim unutarćelijskim parazitima. Naziv virus dolazi od latinske riječi virus, viri = otrovna tvar, toksin.
Virusi uzrokuju mnoge bolesti kod čovjeka, životinja i biljaka, a napadaju i bakterije i druge mikroorganizme. Među ljudskim i životinjskim virusima su osobito poznate boginje, dječija paraliza, gripa, hepatitis, prehlada, bjesnoća, slinavka, šap, žuta groznica, HIV-AIDS, SARS te mnoge druge bolesti. Bolesti uzrokovane virusima nazivaju se zajedničkim imenom viroze, a nauka okoja proučava viruse naziva se virologija.
Definicija virusa prema Fraenkel – Conrat (1988):
Virusi su zarazni uzročnici (čestice) koji sadrže samo jedan tip nukleinske kiseline: DNA ili RNA; njihova je nukleinska kiselina umotana u proteinski omotač izgrađen od jednog ili nekoliko proteina; kod nekih virusa oko proteinskog omotača dolazi složeno građena ovojnica izgrađena od proteina, lipida i ugljikohidrata; virusi su sposobni svoju nukleinsku kiselinu prenositi od jednog do drugog domaćina; sposobni su, dalje, nametnuti ćeliji domaćina svoju genetičku informaciju, zbog čega se mogu koristiti enzimatskim kompleksom domaćina za svoje razmnožavanje; neki virusi mogu svoju nukleinsku kiselinu ugraditi na više ili manje reverzibilan način u DNA domaćina, te tako postati latentni ili perzistentni: neki su od takvih virusa sposobni transformirati genom ćelije domaćina i ometati njezin kontrolni mehanizam rasta, te uzrokovati tumore.
Iako danas postoje brojne definicije virusa, sve one sadrže slijedeće karakteristike:
- virusi sadrže 1 tip nukleinske kiseline (ili RNA ili DNA)
- virusi se repliciraju unutar ćelije
- virusi su obligatni paraziti
3. Historija virologije
Virologija je dosta mlada naučna grana biologije. Prvi virus, virus mozaika duhana - tobaco mosaic virus (TMV), otkriven je 1892. godine. Međutim, iako su virusi otkriveni tek krajem 19. stoljeća, postoje brojni opisi bolesti koje su kolale drevnim civilizacijama. Iz starih kineskih crteža opisana je bolest koja je ličila velikim boginjama, drugi primjeri su reljefi iz starih egipatskih dinastija koji su oslikavali neke bolesti (dječiju paralizu). Jedan od mogućih uzroka propadanja rimskog carstva tumači se epidemijom ospica i velikih boginja. Također, epidemija tipa velikih boginja uzrokovala je umiranje Osteka i na taj način su španci lakše osvojili Južnu Ameriku. Čaki i Aristotel (4. st. p.n.e. ) spominje bjesnoću pasa. Postoje opisi biljnih bolesti iz 8. stoljeća za koje se utvrdilo da su virusnog porijekla. U 16. stoljeću u Nizozemskoj su na cijeni bili tulipani koji su na laticama imali bijele ili žućkaste pruge (virus šarenila cvijeta tulipana).
Prvi napredak u zaštiti od virusnih bolesti učinjen je godinama prije otkrića uzročnika tih bolesti. Gotovo stotinu godina prije Ivanovskijevog otkrića, 1796. godine, britanski liječnik Edward Jenner zapazio je da žene koje muzu krave često obolijevaju od blagog oblika boginja, koje su sa kravljeg vimena prešle na ruke žena. Ta bolest nije imala štetne posljedice za žene, ali je Jenner zapazio da su te žene bile kasnije zaštićene od velikih boginja koje su bile pogubne za čovjeka. Dakle, muzačice su bile na neki način cijepljene ili vakcinirane blagim kravljim boginjama koje su uzrokovale imunost (otpornost) organizma na prave, velike boginje. Jenner je oboljelim osobama ucijepio sadržaj mjehurića iz ruke žene koja je imala kravlje boginje i na taj način ih zaštitio od velikih boginja, te nakon 2 godine objavio rezultate o uspješnoj zaštiti 20 osoba od te bolesti, a u čast krave = vacca, postoji današnji naziv vakcina. 1798. godine načinjeno je cjepivo protiv velikih boginja. 1885. Godine Louis Pasteur je načinio cjepivo protiv bjesnoće, ne znajući da bolest uzrokuje virus.
Otkriće porculanskog filtra omogućilo je otkrivanje čestica koje se danas nazivaju virusi. Prvi čovjek koji je koristio porculanske filtre bio je Adolf Majer, i on je opisao mozaičnu bolest duhana, čiji su se simptomi dosta razlikovali od do tada poznatih gljivičnih bolesti biljaka. Majer je filtrirao sok zaraženih biljaka duhana kroz dvostruki filter papir. Gljivice nije pronašao ni u filtratu ni u filter papiru, i došao je do zaključka da je u filtratu vjerovatno neka bakterija koja je uspjela proći kroz filter papir.
Početkom virologije smatra se 1892. godina. Dimitrij Ivanovski, upoznat sa radovima Majera, filtrira sok bolesnih biljaka i utrljava ih u listove zdravih biljaka. Nakon nekoliko dana na listovima su se počeli pojavljivati simptomi. To je ponavljao i došao do zaključka da u bolesnim biljkama duhana postoji nešto sitnije od bakterija. On je saopštio svoje rezultate u kojima je naveo da je otkrio „filtrabilnu infektivnost“. Prema ‚‚zapadnjačkim’’ knjigama, otkrivačem virusa smatra se Martinus Williem Biejerinku (Bežernik). Međutim, on je svoja istraživanja nastavio na istraživanja Ivanovskog, potvrdio ih i proširio. Također, krajem 19.-tog vijeka postaje jasno da su i uzročnici slinavke i šapa filtrabilni, odnosno ne zadržavaju se na bakteriološkom filtru, a to isto je ustanovljeno i početkom 20.-tog vijeka za uzročnika dječije paralize. Uzročnici su u prvi mah bili nazvani živi otrov, jer istraživači nisu imali predstavu o čemu se tu zapravo radi. Loeffler i Froch (1898.) otkrili su uzročnike odgovorne za bolesti slinavke i šapa. Iste godine kada je Ivanovski otkrio virus, holandski istraživač Bežerink izvodi iste pokuse i objavljuje da je otkrio novi tip infektivnog uzročnika – contagium vivum fluidum. 1915. i 1917.godine dva naučnika su, nezavisno jedan od drugog, otkrili bakteriofage. Godine 1935. W. M. Stanley prvi put uspijeva dobiti virus u čistom stanju, a nakon toga je ubrzo uslijedilo otkriće da u građi TMV-a osim proteina učestvuju i nukleinske kiseline. Godine 1939. zahvaljujući razvoju elektrosnkog mikroskopa, Ruska i sar. načinili su prvi elektronsko-mikroskopski snimak TMV-a.
Savremena istraživanja u virologiji su usko povezana sa razvojem tehnika molekularne biologije. Budući da su jednostavno građeni, virusi su pogodni modeli u fundamentalnim biološkim istraživanjima (reverzna transkriptaza - otkrivena istraživanjem virusa). Virusi su vektori u genetičkom inžinjerstvu. Otkrićem bakteriofaga počela je da se razvija virusologija iz nekoliko razloga - bakterije se vrlo lako gaje i postoji niz metoda koje su omogućile studiranje bakteriofaga u bakterijama, a koje nisu postojale za animalne ili biljne viruse. Animalni i biljni virusi počeli su tek kasnije pomnije da se izučavaju. Danas je situacija takva da su bakteriofagi, animalni i humani virusi najbolje izučeni, dok su biljni virusi ostali najmanje poznata grupa virusa. Danas se dosta izučavaju virusi insekata sa idejom da se koriste u borbi protiv štetnih insekata.
Tabela 1. Najvažnija otkrića u istraživanju virusa
Godina |
Otkriće |
|
|
1892. 1898. 1902. 1911. 1915. 1935.
1937.
1939. |
otkriven prvi virus – virus mozaika duhana – TMV otkriven prvi životinjski virus – virus slinavke i šapa otkriven prvi virus čovjeka – virus žute groznice virusi mogu uzrokovati tumore – Nobelova nagrada otkriveni bakterijski virusi TMV dobiven u čistom stanju, otkrivena mogućnost njegovog kristaliziranja – Nobelova nagrada
otkriven sastav virusa (TMV) : protein i nukleinska kiselina – Nobelova nagrada virus (TMV) prvi put viđen elektronskim mikroskopom – Nobelova nagrada |
1952. |
DNA je genetički materijal bakterijskog virusa i odgovorna je za infektivnost – Nobelova nagrada
|
1956. |
RNA je genetički materijal virusa TMV i odgovorna je za infektivnost |
1970. |
u nekih RNA-virusa otkriven enzim koji prepisuje RNA u DNA (u suprotnom smjeru od onog za kojeg se smatralo da je jedini moguć) – kasnije je dokazano da to nije iznimka, takav se proces odvija i u ćelijama mnogih živih bića –Nobelova nagrada |
4. Opšte karakteristike virusa
Postojanje virusa otkrio je ruski botaničar D. Ivanovski u XIX vijeku, ispitujući mozaičnu bolest duhana, ali je tek otkriće elektronskog mikroskopa omogućilo ispitivanje virusa i upoznavanje njihove građe. Većina virusnih čestica mjeri se nm. Njihove dimenzije kreću se od 20 – 300 nm. Najmanji virusi su biljni satelit virusa nekroze duhana promjera 17 nm, a od animalnih virus slinavke i šapa promjera 20 nm. Od biljnih virusa najveći su klosterovirusi dugi 2000 nm, a od animalnih najveći su pox virusi koji su na granici vidljivosti svjetlosnog mikroskopa. Virusne su komponente molekule sastavljene od vezanih atoma čije vrijednosti izražavamo molekulskom težinom.
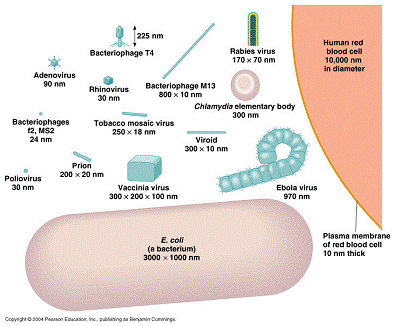
Slika 1. Odnos veličina čestica virusa i bakterijske odnosno ljudske ćelije (eritrocita)
To su aćelijski oblici građeni od proteinskog omotača – kapside i genetičkog materijala – genoma koji je upakovan unutar kapside. Neki virusi mogu imati ovoj – peplos (envelope). Genom virusa su sve nukleinske kiseline sadržane unutar virusne čestice, ili nukleinske kiseline upakovane u populaciji virusnih čestica.
Dok se u ćelijskim organizmima nalaze uvijek obje vrste nukleinskih kiselina DNA i RNA, kod virusa dolazi samo jedna od njih. Nukleinske kiseline su sastavni dio virusnih partikula, a različiti virusi sadrže u čestici različite iznose nukleinskih kiselina. Nukleinske kiseline virusa predstavljaju genom virusa. Genom je skup gena jednog organizma sadržanog u haploidnoj hromosomskoj garnituri. Za razliku od celularnih organizama, kod virusa i RNA može biti nosilac genetičke informacije, pa se razlikuju RNA–virusi i DNA–virusi.
Genom virusa može biti dvolančana ili jednolančana DNA i RNA:
- ds DNA (double stranded – dvolančana DNA)
- ss DNA (single stranded – jednolančana DNA)
- ds RNA (double stranded – dvolančana RNA)
- ss RNA (single stranded – jednolančana RNA)
Tabela 2. Tipovi virusnog genoma
|
dsDNA genom |
ssDNA genom
|
ssRNA genom |
dsRNA genom |
biljni |
Caulimovirusi sa tipičnim članom – virus mozaika karfiola |
Geminiviridae |
većina biljnih virusa ima ss RNK genom: Bromoviridae, Potyviridae, Tombusviridae, Comoviridae i dr. |
Reoviridae |
animalni |
Poxviridae, Herpesviridae, Adenoviridae, Papovaviridae, te virus nuklearne polidroze (virus insekata), Iridoviridae (virus insekata i vertebrata) |
Parvoviridae (Picornavirus) |
Paramyxoviridae (pneumovirusi, virusi zaušnjaka, osipa),Orthomyxoviridae (virus gripa), Rhabdoviridae (virus bjesnila), Retroviridae (virus leukemije HTLV I i HTLV II, HIV),
|
Reoviridae |
bakteriofagi |
T fagi i λ fagi
|
fd (produženi, fleksibilni), Φx 174 |
MS2 |
Q6 |
Genom virusa može biti cjelovita nukleinska kiselina koja može biti različitog oblika – linearna i cirkularna (prstenasta). Genom virusa može biti i fragmentiran tj. podijeljen na 2 do 12 odsječaka. Takvi virusi se zovu virusi sa podjeljenim genomom i kod njih segmenti mogu doći zajedno u jednoj čestici npr. influenca virus (kod njih postoji 8 segmenata RNA) ili reoviride (unutar jedne čestice ima 10 – 12 segmenata) ili segmenti mogu doći u posebnim česticama, i takvi virusi se nazivaju multikomponentni virusi, npr. virus šišavosti duhana u kojem su dva fragmenta genoma upakovana u posebne čestice. Drugi primjer je ss RNA virusa mozaika lucerke čiji je genom građen iz 4 odsječka, a za infekciju je potreban kompletan genom. U jednolančanih RNA virusa (ss RNA) genom se može u procesu replikacije dvojako ponašati, pa prema tome su i označene kao ss RNA+ - pozitivna i ss RNA - – negativna. Virusni genom je RNA+ kada se u procesu replikacije ponaša kao iRNA (informaciona RNA). RNA+ nakon oslobađanja proteina kapside u ćeliji se veže za poliribosome i započinje sintezu ranih proteina tj. enzima koji će katalizirati sintezu virusnih genoma potomaka. RNA- genom imaju oni virusi čiji je genom komplementaran informacionoj RNA, tj. za aktiviranje genoma neophodan je enzim RNA transkritptaza ugrađen u kapsidi koji će omogućiti transkripciju RNA- u iRNA tj. RNA+. Najviše biljnih virusa ima RNA+.
Posebno su interesantni RNA-DNA virusi zbog toga što posjeduju ili RNA kao genetički materijal, ali se razmnožavaju preko DNA intermedijera, ili posjeduju DNA kao genetički materijal, ali se replikacija vrši preko RNA intermedijera. Prvu grupu nazivamo retrovirusi i oni imaju jednolančanu RNA kao genom. Retrovirusi čine posebnu grupu virusa. Sadrže RNA, a od ostalih virusa se razlikuju po tome što pomoću vlastitog enzima svoju genetičku uputu prevode u DNA. Pripadaju im virsi koji su uzročnici najtežih bolesti, umora i AIDS-a. Drugu grupu čine hepadna virusi koji imaju dvolančanu DNA kao genetički materijal
Danas je poznata nukleotidna sekvenca tj. primarna struktura mnogih virusa. Prvi puta je sekvenciran kopletan genom 1976. godine, i to bakteriofaga MS2 , koji se sastoji od 3596 nukleotida. Genom faga MS2 sadrži tri gena, jedan nosi informaciju za A protein, drugi gen za strukturni protein kapside, a treći gen za RNA replikazu. Interesantna je struktura jednolančanog genoma gemini virusa (tj. blizanci) koja ima dvije molekule DNA slične dužine, ali različite sekvence. DNA I ima 2779 nukleotida, a DNA II je duga 2724 nukleotida i svaka partikula blizanaca nosi samo jednu molekulu genoma, pa se takav genom naziva bipartitni (dvodijelni) genom. Sekvenca genoma virusa HIV prvi put je određena 1985. godine. Predstavljena je sa dvije kopije ss RNA, tj. virion nosi dvije kopije genoma, a nije poznato koja od njih je funkcionalna. Virusne nukleinske kiseline sadrže uobičajne baze (nukleotidi su građeni od purinske i pirimidinske baze, šećera riboze u RNA i dezoksirboze u DNA i ostataka fosforne kiseline), a neki bakteriofagi umjesto citozina imaju 5 – hidroksi metil citozin tj. 5 hidroksi – metil uracil, a umjesto timina 5 – hidroksipenil uracil.

Slika 2. Građa virusne čestice - viriona

Slika 3. Građa virusa: a) virus građen samo od nukleokapside, tzv. „goli virus“;
b) forma virusa sa ovojnicom
Za infekcioznost virusa značajna je kapsida – proteinska ljuska koja obavija nukleinsku kiselinu. Kapsida je građena od proteinskih podjedinica = protomere, koje mogu biti kopije jednog proteina ili kapsidu gradi uglavnom manji broj različitih proteina, što govori o ekonomici virusnog genoma (za večći broj proteina bio bi potreban i veći broj gena koje se ne bi mogli upakovati unutar kapside). Jednom nastale protomere se udružuju međusobno tj. specifično se povezuju tvoreći kapsidu. U tvorbi kapside nema pomoći iz vana pa se proces njenog nastanka naziva samoudruživanje ili samosastavljanje (slično nastanku bakterijskog flageluma). Zahvaljujući strukturi i organizaciji svojih podjedinica, kapsida izvanredno štiti virusni genom, pomaže njegovom prenosu među ćelijama domaćina, nosilac je antigena. Građena je od kapsomera i nosilac je virusnih antigena. Kapsomere se međusobno spajaju u najraznovrsnije geometrijske oblike koji daju virusima izgled štapića, poliedara, metka, rakete, kristala itd. Mnogi virusi preko kapsida na svojoj površini imaju virusni omotač koji sadrži peptide i polisaharide, a neki i lipide. Iz nje strše glikoproteinski izdanci kojima se virus prihvata za ćeliju primatelja. Takvi su npr. virusi gripe, bjesnoće i HIV-a. Životinjski virusi prilikom izlaska iz inficirane ćelije dobijaju omotač od izmijenjenih ćelijskih membrana. Jedan dio proteina izgrađuje kapsidu (građevni proteini) i određuju oblik virusa : kod produženih oblik valjka, a kod poliedričnih oblik pravilnog poliedra. Drugi dio proteina su virusni enzimi (funkcionalni enzimi). Složeniji virusi koji sadrže ovojnicu sadrže više vrsta proteina. Nukleinska kiselina čini 5–40 % građe virusne čestice, dok ostatak od 60-95 % ide na proteine. U česticama biljnih virusa mogu se naći i anorganski i organski kationi, te znatne količine vode.
Prema morfologiji kapside na osnovu elektronsko-mikrsokopskih pretraga virusi se mogu klasificirati u nekoliko morfoloških tipova:
- produženi virusi (izometrični ili poliedrični),
- polimorfni,
- kompleksno građeni virusi,
- multikomponentni virusi.
Produženi virusi liče dugom štapiću, a mogu biti kruti ili fleksibilni. Oblik krutog štapića ima TMV ili bakteriofag MB, a fleksibilnog štapića poti virusi ili npr. closterovirus.
Izometrični (poliedrični) virusi su više ili manje loptastog oblika i njihove dimenzije mjere se prečnikom, tu spadaju različiti virusi – HIV, rota virus, herpes virus. Od izometričnih virusa treba spomenuti gemini viruse ili blizance čije se čestice javljaju u parovima.
Polimorfni virusi pored kapside posjeduju i ovojnicu. Postoje produženi i izometrični, a imaju pleomorfan oblik iz razloga što im ovojnica nije rigidna tj. kruta. Primjer za produžene viruse sa ovojnicom jeste virus gripe ili influence ili ebola virus, dok su herpes virus ili HIV primjer za izometrične viruse sa ovojnicom.
U skupini komleksno građenih virusa spadaju neki bakteriofagi i pox virusi. Bakteriofagi su specifičnog oblika. Najčešće su građeni od glave i repa, no ima ih i građenih samo od glave tako da izgledaju kao izometrični virusi. Obično im je genom dlDNA. U glavi se nalazi nukleinska kiselina koju ubacuju u bakterijsku ćeliju. Imaju litički i lizogeni ciklus. Litički je onaj kod kojeg odmah nakon umnožavanja virusa dolazi do lize ćelije. Kod lizogenog se ciklusa genom faga ugrađuje u genom bakterije (profag) i replicira se s njim u diobama bakterijske ćelije. Do lize dolazi nakon dužeg vremena i to u određenim uslovima. Vakcinija virusi je virus kravljih boginja, spada među animalne viruse a ima oblik lepne. Rabdovirusi imaju poseban oblik , oblik puščanog zrna, čiji je jedan kraj ravan a drugi zaobljen.
Od multikomponentnih virusa poznat je virus mozaika lucerke i virus šuštavosti duhana. Virus mozaika lucerke u populaciji ima 5 tipova čestica, 4 baciloformne i jedna okrugla, a kod virusa sušenosti duhana dvije.

Slika 4. Osnovni oblici i građa biljnih virusa: a) nitasti, b) štapičasti, b-1 i b-2) pogled na građu kapside i srži i c i c-1) baciliformni, d i d-1) izometrični (okruglasti oblici) i e) čestica geminivirusa od dvije spojene čestice, NK- kukleinska kiselina, PP- proteinska podjedinica

Slika 5. Bakteriofag
Iako su virusi obligatni paraziti slično nekim bakterijama kao što su rikecije, hlamidije i mikoplazme, oni se od navedenih intracelularnih parazita bitno razlikuju u pogledu strukturne organizacije, hemijskog sastava, fiziološki, domaćina i načina parazitizma.
- strukturno
Za razliku od njih, virusi su acelularni oblici i ne pripadaju prokariotima. Vrlo su jednostavne građe i dugo se mislilo da su najjednostavnije građeni patogeni, danas su poznati oblici građeni jednostavnije od virusa (viroidi, sateliti virusa, virusne RNA, prioni). Zbog opšte građe virusa uopšte je prihvaćen naziv za tijelo virusa – virusna čestica ili partikula.
- hemijski
Virusi i po hemijskoj građi pokazuju osebujnost i razlikuju se od celularnih patogena. Svi celularni organizmi imaju obje vrste nukleinskih kiselina, međutim virusi imaju samo jednu, ili RNA ili DNA.
- fiziološki
Virusi su paraziti koji nemaju vlastiti metabolizam, te ne mogu rasti i razmnožavati se kao ostali organizmi. Dok se mikroorganizmi razmnožavaju diobom ćelija, umnožavanje virusa, tj. propagacija – multiplikacija, provodi se genetički progamiranom sintezom virusnih komponenti i njihovim udruživanjem. Nakon prodiranja u ćeliju, virusna nukleinska kiselina se oslobađa proteinskog omotača i koristeći ćelijski sintetički aparat sintetizira nove virusne partikule (prisiljava ćeliju domaćina).
- domaćin
Opšta karakteristika parazita je da su njihovi domaćini biljke, životinje ili čovjek, međutim neki virusni patogeni biljaka razvijaju se na domaćinima i biljnog i životinjskog svijeta. Takvi virusi nazivaju se perzistentnim, propagativnim, odnosno cirkulatornim virusima.
- parazitizam
Svi intracelularni paraziti koriste ćeliju domaćina kao izvor potrebnih materija koje uključuju u sopstveni metabolizam i na taj način obezbjeđuju vlastite životne funkcije. Naprotiv, virusi svojim prisustvom mijenjaju metabolizam ćelije i usmjeravaju ga u pravcu sinteze sopstvenih virusnih partikula, što može imati različite posljedice za ćeliju domaćina, pa čak dovesti i do fatalnih posljedica. Parazitizam virusa je poistvojećen sa multiplikacijom virusa, što predstavlja novi kvalitativni oblik parazitizma.
5. Simetrija virusnih čestica
Morfološka organizacija virusne čestice određena je strukturom njene kapside tj. rasporedom njenih podjedinica. Postoje 2 tipa virusne simetrije:
- spiralna i
- kubična
Virusi spiralne simetrije su produženi, imaju spiralan raspored podjedinica kapside koje omotavaju centralno položenu nukleinsku kiselinu u obliku spirale (heliksa). Prema obliku kapside, takve virusne čestice imaju valjkast ili cilindričan oblik. Većina biljnih virusa ima takvu simetriju, npr. TMV, karla virusi, postero virusi, poti virusi i dr. Ne postoje animalni virusi ove simetrije, odnosno spiralni, a da nemaju ovojnicu.
Kubična simetrija se nalazi u poliedričnih virusa čije je kapsida izometrična. Najpoznatiji je ikozaiedar koji susrećemo kod životinjskih virusa (herpes virus). Kod takvih virusa površina viriona je sastavljena od 20 istostraničnih trogulova i ima 12 vrhova i 30 ivica. Kroz središte tog geometrijskog tijela mogu se povući 3 ose rotacijske simetrije:
1. jedna kroz svih 12 vrhova
2. druga kroz središte od 20 ravni trokuta
3. treća sredinom svakog od 30 bridova (ivica)
Rotacijom jednom od 3 ose simetrije slika ikozaiedra se pojavljuje redom 5, kroz drugu 3 i kroz treću 2 puta. Po obliku kapside virusi imaju poliedričnu formu. Budući da proteinske molekule nemaju oblik istostraničnog trougla, u najjednostavnijih kapsida su dovoljne 3 identične PP za izradu 1 trokuta, a kod 20 – straničnog 30 PP. Viši stupnjevi organizacije ikozaiedra nastaju daljom diobom površinskog trokuta, to je tzv. triangulacijski broj koji označava koliko je manjih trokuta nastalo od izvornog trokuta. Proteinske ili strukturne podjedinice izometričnih virusa mogu se skupljati u veće tvorbe koje su sastavljene od više protomera i nazivaju se kapsomere – morfološke podjedinice. Npr. 60 pomenutih protomera može se u kapsidi nekih virusa rasporediti tako da se po 5 njih skupi na svaku od 12 vrhova ikozaiedra. Takvu građu ima npr. čestica satelitskog virusa uz virus nekroze duhana koja ima 12 pentamera. Kompleksno građeni virusi bakteriofaga imaju glavu kubične, a rep spiralne simetrije. Mnogi virusi imaju kompleksnu ovojnicu koja okružuje nukleo–kapsidu. Ovojnica je građena od lipidnog dvosloja u koju su uloženi glikoproteidi. Lipidi potiču od membrane ćelije domaćina dok glikoproteine kodira virusni genom. Simetrija virusa sa ovojnicom određuje se na osnovu nukleokapside koja se nalazi unutar ovojnice. U viruse sa ovojnicom spadaju poznati virusi iz familije Ortomiksoviride, Paramiksoviride, Rabdoviride, a kubične simetrije fam. Retroviride u koju spada HIV.
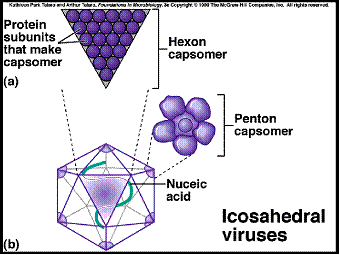
Slika 6. Kubična simetrija virusne čestice

Slika 7. Spiralna simetrija virusne čestice
6. Subviralni patogeni – subviralne infektivne molekule
Viroidi. Predstavljaju infektivne čestice manje od virusa. Gole su jer ne posjeduju kapsidu. To su jednolančane kružne molekule RNA koje su infektivne i uzrokuju bolesti viših biljaka. Ne upravljaju sintezom proteina jer su suviše mali. Viroidna RNA dosta je stabilna i neobično infekciozna; izračunato je da u 1g ukupne RNA, koja je izolirana iz biljke inficirane viroidom, ako se razrijediti u gotovo 2 miliona litara vode takva otopina ostaje infekciozna. Otkriveni su 1971. god., a otkrio ih je Diner koji je tvrdio da su viroidi čista RNA, te im je on dao naziv viroid. Najpoznatiji je viroid vretenastog gomolja.
Prioni. Drugi tip patogena su prioni. Otkrio ih je Stanley B. Prusiner 1996. god, za šta je dobio Nobelovu nagradu. Prioni su proteinske infektivne čestice, to su glikoproteini koji mogu biti infektivni. Takvi proteini nalaze se u plasmamembrani normalnih ćelija i još uvijek im se ne zna uloga. Prisutni su ćelijama nervnog tkiva i označeni su kao PrPc, a oni koji uzrokuju bolest označeni su kao PrPsc (scscrapie). Prioni su uzročnici bolesti ludih krava [bovine spongiform encephalopathy (BSE)], zatim nasljedne krojcfeld – jakobsonove bolesti (Creutzfeldt-Jakob disease) i mnogih drugih bolesti.
Sateliti virusa. Neki biljni virusi ne mogu se razmnožavati samostalno, zbog toga u prirodi uz njih uvijek dolazi virus pomagač. Odnos između satelitnog virusa i virusa pomagača (helper virusa) strogo je specifičan. Najpoznatiji je satelitni virus koji dolazi uz virus nekroze duhana. Satelitni virusi se ne mogu samostalno razmnožavati, ali mogu stvoriti vlastiti proteinski omotač, tj. njihova RNA dolazi u zasebnim česticama.
Satelitne RNA. Postoji i druga vrsta satelita koja nema vlastitu kapsidu tj. u virusnim česticama su nađene strane ssRNK koje nisu dio virusnog genoma. One se također ne mogu samostalno replicirati, tj. bez pomoći virusa pomagača. To su satelitne RNA koje ne mogu stvarati vlastiti proteinski omotač več dolaze unutar kapside virusa pomagača npr. satelitna RNA virusa mozaika krastavca.
7. Evolucija virusa
Pitanje o porijeklu virusa i njihovoj prirodi je predmet mnogobrojnih istraživanja naučnika. Teorije o porijeklu virusa uglavnom se svode na tri osnovne koncepcije.
- Po prvom gledištu virusi predstavljaju aćelijski oblik živih bića i označeni su zajedničkim imenom protobionti. Zastupnici ovog gledišta smatraju ih potomcima prvobitnih organizama koji su se u davnoj prošlosti pojavili na Zemlji.
- Po drugom gledištu, virusi predstavljaju regresivne oblike bakterija koje su izgubile ćelijsku građu uslijed dugotrajne prilagođenosti na parazitski način života u ćelijama živih bića.
- Najzad, treće gledište svodi se na to da virusi uopšte ne pripadaju živim bićima. Ovo tvrdnja se zasniva na činjenici da mnogi virusi obrazuju kristale, slične mineralima, neorganske prirode. Pojava kristalizacije naročito je karakteristična za sitne viruse.
Nauka je do sada prikupila veliki broj dokaza koji potvrđuju shvatanje da se život može manifestovati ne samo u ćelijskom, već i u aćelijskom obliku organizacije, kakvu predstavljaju
virusi. Najinteresantniji problem koji biologija pokušava da riješi je prelaz od neživog ka živom. Danas se zna da postoje i prostije čestice od virusa - viroidi, tj. kratki segmenti RNA koji mogu da se repliciraju u živim ćelijama izazivajući oboljenja (npr. bolest češanja kod ovaca). Još interesantnija je pojava priona - proteina male molekulske težine koji se samostalno razmnožavaju i izazivaju bolest ludih krava. Ova otkrića pomjeraju saznanja nauke i mijenjaju kriterijume o neživom i živom.
Danas, najprivaćenija hipoteza o nastanku virusa je ta da su virusi nastali iz dijelova ćelije domaćina - to su fragmenti nukleinske kiseline, lutajući geni“ koji su izbjegli kontroli ćelije i počeli se autonomno razmnožavati.
8. Klasifikacija virusa
Za razliku od klasifikacije eukariota, klasifikacija virusa se često mijenjala, dopunjavala i još uvijek nije u potpunosti zadovoljavajuća. Naučna klasifikacija virusa je složen posao, jer je viruse teško grupisati samo po jednom kriterijumu. U ranijim fazama taksonomije virusa, klinička i patogena svojstava, kao i ekološki karakteri i prijenosnici virusa, bila su glavna obilježja koja su se koristila za klasifikaciju virusa. Virusi su se pokušavali svrstati u grupe na osnovu tipa bolesti npr. virus mišje groznice, virus hepatitisa, potom prema domaćinu: animalni, biljni, bakterijski virus ili prema ciljnim organskim sistemima : virus probavnog sistema, virus disajnog sistema itd. Bilo je klasifikacija prema vektoru, npr. arbovirus (prenosioci arthropode). U imenovanju virusa korišteni su neki termini koji opisuju osobine virusa npr. piko RNA (piko – mali, RNAtip nukleinske kiseline), toga virus (toga – omotač), papova virus (od početnih riječi pa – papiloma, po – poliom, va – vacuola), tj. od najvažnijih patoloških simptoma, retrovirusi – prema reverznoj trankriptazi, koksaki virusi – prema mjestu u SAD. Biljni virusi najčešće dobijaju ime po biljci domaćinu i simptomima koje uzrokuju, npr. TMV.
Otkrićem elektronskog mikroskopa i njegovom širokom primjenom, morfologija virusne čestice postaje najvažnija karakteristika za klasifikaciju virusa. Iako je većina virusa vidljiva elektronskim mikroskopom, danas se teži tome da se upozna hemijska i genomska struktura virusne čestice, kompleks simptoma bolesti koje izazivaju u svojim domaćinima, geografska distribucija, kao i vektori koji ih prenose. Stabilnost virusne čestice (uticaj promjene pH i temperature, izloženost otapalima lipida, deterdžentima i sl.) i njeni antigeni (određeni različitim serološkim metodama), također predstavljaju bitne kriterije za taksonomiju virusa. Sekvencioniranje genoma virusa se sve češće obavlja vrlo rano u identifikacijskim protokolima, čak i u zemljama u razvoju. Međutim, u većini slučajeva pri svrstavanju virusa u specifične taksonomske kategorije, morfologija virusne čestice i rezultati seroloških testova i dalje ostaju važni kriteriji za identifikaciju nepoznatih virusa. U nekim slučajevima, analiza sekvence ne može identificirati virusnu česticu do nivoa potrebnog za preciznu kliničku dijagnozu, pa se tada koriste serološki testovi za razlikovanje blisko povezanih serotipova (npr. razlikovanje između japanskog encefalitis virusa, Murray Valley encefalitis virusa i West Nile virusa).
Danas, klasifikacije se najčešće vrše prema :
- tipu nukleinske kiseline na RNA i DNA - na jednostruko ili dvostruko lančanu;
- organizmima (ćelijama) u kojima parazitiraju - na viruse bakterija (bakteriofagi), virusebiljaka, viruse životinja i viruse čovjeka;
- veličini i tipu simetrije, broju kapsomera i prisutnosti membrana;
- osjetljivosti na fizičke i hemijske faktore, naročito prema etru;
- imunološkim svojstvima;
- načinima prenosa;
- tropizmu prema domaćinu, tkivu i ćeliji;
- patologiji;
- simptomatologiji.
U nastojanju da se stvori jedinstvena taksonomija virusa, u Moskvi je 1966. god. osnovan Međunarodni odbor za nomenklaturu virusa, a 1981. god. odbor je predložio klasifikaciju prema kojoj se virusi na temelju osobina svrstavaju u familije sa sufiksom -viridae, podfamilije -virinae, rod -virus i vrste. Još uvijek je veliki broj novootkrivenih virusa nesvrstanih zbog nepotpunih podataka o njihovim virološkim, fizičkim i hemijskim osobinama.
Danas, primarni kriteriji koji se koriste za razlikovanje redova, familija
i rodova virusa
su slijedeći:
- vrsta i organizacija virusnog genoma
- strategija virusne replikacije
- struktura viriona (virusne čestice)
9. Historija taksonomije virusa
Najraniji eksperimenti koji uključuju viruse odvojili su viruse od drugih mikroba koji se mogu vidjeti svjetlosnim mikroskopom i koji se mogu uzgajati na prilično jednostavnim medijima. U eksperimentima koji su doveli do prvog otkrića virusa, Beijerinck i Ivanovski (virus mozaika duhana), Loeffler i Frosch (slinavka i šapa virus) i Reed i Carroll (virus žute groznice) na kraju stoljeća, mjerena je samo jedna fizičko-hemijska karakteristika, filtrabilna infektivnost virusnih čestica. Niti jedna druga fizička osobina nije mjerena u to vrijeme, i većina proučavanja virusa bila je usredotočena na njihove sposobnosti da uzrokuju infekcije i bolesti. Najranija nastojanja da se izvrši klasifikacija virusa, dakle, temeljla su se na zajedničkim svojstvima patogena, zajedničkom tropizmu organizama, te zajedničkim ekološkim karakteristikama i načinu prijenosa. Na primjer, patogeni virusi koji uzrokuju hepatitis (npr. virus hepatitisa, virus hepatitisa B, virus hepatitisa C, virus žute groznice i Rift Valley virus groznice), označeni su jednim imenom "virusima hepatitisa". Biljni virusi koji uzrokuju mozaike (npr. virus mozaika cvjetače, virus mozaika ljulja, virus mozaika lucerne, virus mozaika duhana), također su svrstani u istu grupu virusa "mozaik virusa."Iako su prva proučavanja virusa počela na prijelazu stoljeća, do 1930. god. nisu postojali dokazi o građi i sastavu virusne čestice. To je potaknulo Bawden (1941., 1950.) da viruse klasificira na osnovu zajedničkih osobina virusne čestice. Među prvim taksonomskim grupama izgrađenim na toj osnovi bile su grupe herpesvirusa (Andrewes, 1954.), grupe myxovirusa (Andrewes, Bang i Burnet, 1955.), grupe poxvirusa (Fenner i Burnet, 1957.), te nekoliko grupa biljnih virusa sa virusnom česticom u obliku štapića ili niti (Brandes i Wetter, 1959.).
U 1950.-im i 1960.-im došlo je do eksplozije u otkrivanju novih virusa. Potaknuti velikom količinom informacija, nekoliko pojedinaca i odbori samostalno kreiraju napredne klasifikacijske sheme, što je dovelo do potpune pomutnje u tadašnjoj klasifikaciji virusa. Tada je već postalo jasno da je klasifikacija i nomenklatura virusa vrlo složen i zahtjevan posao.
Tokom godina bilo je mnogo pogrešaka i duplikacija prilikom imenovanja i klasifikacije virusa. Mnogi virusi izolirani od ljudskih, životinjskih i biljnih uzoraka izolirani su u različitim laboratorijama, dobili različite nazive i samim tim i različita mjesta na listi virusa. Razlozi za ove i slične probleme su: (1) neadekvatna karakterizacija i opis virusa od strane onih koji su ih izolirali, i (2) neadekvatan pregled podataka međunarodnih specijalnih grupa. Iako su danas takvi problemi rijetkost, i dalje se klasifikaciji virusa pridaje posebna pažnja, jer pogreške mogu imati ozbiljnih posljedica.
10. Sistemi klasifikacije virusa
- Holms-ov sistem klasifikacije (1948. godina)
Prati Linneov sistem binarne nomenklature. Prema ovoj klasifikaciji, virusi su podijeljeni u tri grupe unutar reda virales :
• Grupa I : phaginae
(napada bakterije)
• Grupa II : phytophaginae (napada biljke)
• Grupa III : zoophaginase (napada životinje)
- LHT sistem ili klasični sistem za klasifikaciju virusa (1962.
godina)
Lwoff, Robert W. Horne i Paul Tournier koriste Linneov hijerarhijski
sistem za klasifikaciju virusa. Klasifikacija se temelji na nivou odjeljka,
klase, reda, familije, roda i vrste.
• Virusi su grupisani prema njihovim zajedničkim svojstvima (ne
svojstvima domaćina)
• hemijska i fizička svojstva (npr. nukleinske kiseline (DNA ili RNA),
simetrija (spiralna
ili kubična), prisustvo i odsustvo ovojnice itd.
- Baltimore-ova klasifikacija (David Baltimor, 1970. godina)
S obzirom na način sinteze iRNA, američki virolog David Baltimore (otkrio reverznu transkriptazu) svrstao je sve viruse u 6 grupa:
- ds DNA virusi
- ss DNA virusi
- ds RNA virusi
- RNA +
- RNA -
- reverzno transkribirajući virusi
Baltimore je iRNA označio kao (+)RNA, a sve virusne nukleinske kiseline koje su joj komplementarne označene su znakom (-).
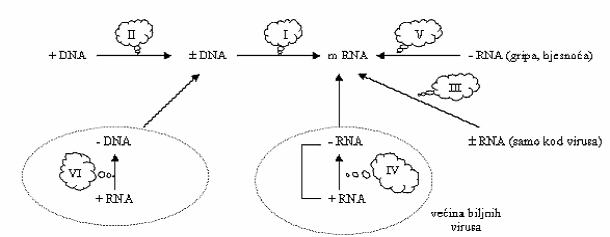
Slika 8. Baltimore-ova klasifikacija virusa
I: dlDNA-virusi- animalni virusi, neki biljni virusi, npr. vir. mozaika cvjetače
(= dobar prirodni vektor za ubacivanje strane, rekombinantne DNA u genom biljaka.
II: jlDNA-virusi- ova pojava postoji samo kod virusa, npr. geminivirus
III: dlRNA-virusi- npr. virus mozaika salate
IV: (+)jlRNA-virusi - većina biljnih virusa
V: (-)jlRNA-virusi - npr. virus bjesnoće, gripe, neki biljni virusi
VI: jlRNA- retrovirusi (od biljnih samo virus mozaika cvjetače
- Casjens-ov i Kings-ov klasifikacijski sistem (1975. godina)
Ovaj sistem
klasifikacije virusa temelji se na četiri karakteristike virusa
:
nukleinske kiseline, prisustvo omotnica, simetrija virusne čestice i mjesto
pojavljivanja
- Međunarodni komitet za taksonomiju virusa - ICTV = International Committee on Taxonomy of Viruses (1973. godina)
11. Međunarodni komitet za taksonomiju virusa (ICTV)
Međunarodni kongres za mikrobiologiju 1966. u Moskvi osnovao je Međunarodni komitet za nomenklaturu virusa (ICNV). Već tada virologisti su osjećali potrebu za jedinstvenom, univerzalnom taksonomskom shemom virusa. Bilo je teško i sporo razvrstati stotine virusa izoliranih od ljudi, životinja, biljaka, beskičmenjaka i bakterija u jedan sistem, a onda taj sistem odvojiti od ostalih bioloških grupa. Lwoff, Horne i Tournier (1962.) su predložili sveobuhvatnu shemu za klasifikaciju virusa u pododjeljke, klase, redove, podredove i familije. Hijerarhijska podjela virusa na ove glavne taksonomske grupe, samovoljno i monotetički, temeljila se na tipu nukleinske kiseline, simetriji kapside, prisustvu ili odsustvu omotnice itd. Mnogi virolozi se nisu složili sa ovakvom podjelom, protiveći se samovolji u odlučivanju koje karakteristike virusne čestice trebaju imati relativnu važnost pri klasifikaciji virusa. Tvrdili su da se još uvijek nedovoljno zna o karakteristikama većine virusa da bi ovakva hijerahijska podjela bila prihvaćena. Alternativni prijedlog 1966. dali su Gibbs i sar. U tom sistemu, podjela je izvršena na osnovu više kriterija (politetički kriteriji). Sistem je predstavljen korištenjem "kriptograma" (kodirani zapisi osam virusnih karakteristika). Ova rana nastojanja da se izvrši klasifikacija virusa, dovela su do razvoja sistema univerzalne taksonomije u 1970.-tim godimama, koja se vremenom mijenjala i održala sve do danas. Međunarodni komitet za nomenklaturu virusa (ICNV) postao je Međunarodni komitet za taksonomiju virusa (ICTV) u 1973. godini. Danas, ICTV djeluje pod pokroviteljstvom Odjela za virologiju Međunarodne unije mikrobioloških društava. ICTV ima šest pododbora, 45 studijskih grupa i preko 400 virologa. Prema univerzalnoj shemi ICTV-a, osobine virusne čestice se smatraju važnim kriterijem za podjelu odjeljaka na familije, u nekim slučajevima i na podfamilije, i rodove. U početku, klasifikacija virusa po ovoj shemi nije dijelila viruse na taksonomske grupe iznad familije. Takson vrsta u klasifikaciju virusa uveden je tokom 1990.-ih i tek tada je postalo jasnije da se familije i rodovi najbolje mogu definirati monotetički (ili na osnovu samo nekoliko karakteristika), a vrste politetički. Na sjednici u Meksiku 1970. godine, ICTV je odobrio prve dvije familije i 24 rodova. U to vrijeme, 16 grupa biljnih virusa su također imenovane. Od tada, ICTV je objavio osam izvješća pod naslovom Klasifikacija i nomenklatura virusa (1971., 1976., 1979., 1982., 1991., 1995., 2000., 2009.). Osmi izvještaj ICTV evidentirao je univerzalnu taksonomsku shemu virusa koja se sastoji od šest redova, 86 familija, 19 podfamilija, 348 rodova i 2290 vrsta. Sistem i dalje sadrži stotine „neraspoređenih“ virusa, uglavnom zbog nedostatka podataka.Međunarodni komitet za taksonomiju virusa (ICTV) 1999. godine uveo je sistem decimalnih kodova i omogućio kompletnije identifikovanje i tačnije određivanje taksonomskog statusa virusa.
12. Međunarodna baza podataka za taksonomiju i klasifikaciju virusa - ICTVdB
ICTVdB je osnovana na Australian National University, a danas je bazirana na Sveučilištu Columbia. ICTVdB koristi DELTA sistem (programski jezik za opisivanje taksonomije), koji je usvojen kao svjetski standard za razmjenu podataka u taksnomiji. Posebnost ovog programa je mogućnost pohrane ogromnog broja različitih podataka (tekst, tabele, slike, komentari i sl.) i prevod podataka za tradicionalna izvješća i web izdanja. Još jedna prednost ICTVdB je mogućnost on-line unosa podataka, čime se omogućava praćenje novih informacija, od molekularnih svojstava, pa sve do geografske distribucije virusa i domaćina koje inficiraju. ICTVdB se temelji prvenstveno na hemijskim karakteristikama virusa, njihovom genomskom tipu, replikaciji nukleinskih kiselina, bolestima koje izazivaju, vektorima koji ih prenose, domaćinima te geografskoj distribuciji.13. Međunarodni kodeks klasifikacije i nomenklature virusa
Osnovni ciljevi Kodeksa klasifikacije i nomenklature ICTV su:
- razviti međunarodno dogovorenu taksonomiju virusa,
- razviti međunarodno dogovorena imena za svojte virusa, uključujući i vrste i subvirusne čestice,
- da međunarodna zajednica virologa zajedno donosi taksonomske odluke.
Osnovni principi nomenklature virusa su:
- stabilnost
- izbjegavanje ili odbijanje korištenja imena koja bi mogla dovesti do pogreške ili zabune
- nepotrebno stvaranje imena
Nomenklatura
virusa i subvirusnih čestica je neovisna od ostalih bioloških nomenklatura
i ima status „iznimke“ u predloženom Međunarodnom kodeksu
bionomenklature (BioCode). Primjena imena taksona utvrđuje se, eksplicitno
ili implicitno, pomoću nomenklature vrste.
Ime taksona nema službeni status sve dok ga ne odobri Međunarodni komitet
za taksonomiju virusa ( ICTV).
13.1. Pravila klasifikacije i nomenklature virusa
Klasifikacija i nomenklatura virusa mora biti internacionalna i univerzalno se primijenjivati na sve viruse. Univerzalni klasifikacijski sistem virusa raspoređen je u slijedeće taksonomske kategorije : red, familija, podfamilija, rod i vrsta. Prilikom klasifikacije nije obavezno koristiti sve nivoe taksonomske hijerarhije. Primarna klasifikacija virusa je u vrste. Većina vrsta su razvrstane u rodove i većina rodova su svrstani u familije. Vrste koje nisu klasificirane u rodu su "neraspoređene" u familiji, rodovi koji nisu klasificirani u familiji imaju status "neraspoređen" (ponekad se nazivaju "plutajući"). Neke familije se zajedno klasificiraju u redove, ali za mnoge, familija je najviši takson u upotrebi pri klasifikaciji virusa. Također, familije nisu nužno svrstane u podfamilije. Ovaj takson se koristi samo kada je potrebno riješiti neke složene hijerarhijske probleme.
Primjeri pune klasifikacije nekog negativnog soja RNA virusa su: - (1) vrsta Mumps virus; rod Rubulavirus, podfamilija Paramyxovirinae, familija Paramyxoviridae, red Mononegavirales, (2) vrsta Rice stripe virus; roda Tenuivirus.
13.2. Ograničenja pri klasifikaciji
Tek kada je reprezentativni član virusa dovoljno dobro okarakteriziran i opisan u literaturi i nedvosmisleno se razlikuje od drugih sličnih svojti, takson će biti priznat (utemeljen, imenovan). Ako je klasifikacija vrste u familiji jasna, ali je neizvjesno u koji rod treba klasificirati vrstu, takva vrsta se klasificira kao „neraspoređena“ vrsta te familije. Vrsta može biti klasificirana kao „neraspoređen član“ familije, ako rod nije otkriven (određen). Taksoni iznad vrste moraju biti odobreni prije nego im se dodijeli ime.
13.3. Pravila o imenovanju taksona
Predložena imena za taksone su "validna" ako su u skladu s Pravilima utvrđenim kodeksom i ako se odnose na priznate taksone. Validna imena se smatraju "prihvaćenim imenima" ako su evidentirana kao međunarodno odobrena imena u 8. ICTV izvještaju ili su naknadno postala "prihvaćena imena" glasovanjem članova Međunarodnog komiteta o odobrenju taksonomskog prijedloga. Važećim imenom se smatra ono ime koje je objavljeno i povezano sa opisnim materijalom i koje zadovoljava pravila Kodeksa. Prihvaćena imena se nalaze u "Indeksu imena" ICTV-a. Stabilna nomenklatura je jedan od glavnih ciljeva taksonomije, te se promjena imena koje su prihvatili članovi Odbora razmatra samo u iznimnim okolnostima, ili ako dođe do ozbiljnih sukoba s pravilima Kodeksa.Pri imenovanju taksona i virusa nema prioriteta. Pri osmišljavanju imena za nove svojte ne smiju se koristiti imena osoba. Nova imena taksona ne smiju se kreirati usvajanjem ličnog imena, dodajući formalni završetak svojte na ime osobe ili koristeći dio ličnog imena kao osnovu za stvaranje imena. Imena za taksone trebaju biti jednostavna za korištenje i laka za pamćenje. Kratka imena su poželjnija, a broj slogova treba svesti na najmanju moguću mjeru.
Indeksi, eksponenti, crtice, kose crte i grčka slova ne smiju se koristiti u osmišljavanju novih imena. Novo odobrena imena ne smiju se ponavljati. Nova imena se odabiru tako da ona nisu blisko slična imenima koja su trenutno u upotrebi ili su se upotrebljavala u nedavnoj prošlosti. Na primjer, postojanje roda Iridovirus znači da su oblici novih imena kao što su "irodovirus" ili "iridivirus" neprihvatljivi, jer ih je lako pomiješati s odobrenim imenom. Zbunjenost može biti između imena vrste i roda koji završavaju na "virus". Tako, na primjer, rod kojem pripada vrsta "Omega virus" ne može se zvati "Omegavirus", jer bi vrlo lako moglo doći do zamjene vrste i roda. Skraćenice mogu biti prihvaćene kao imena taksona, pod uslovom da su te skraćenice značajne za određeno polje u virologiji. Skraćenice čine imena slova i/ili kombinacije slova preuzetih iz složenica. Ime roda Comovirus je skraćenica nastala od matične riječi "Co-" crni grah i od "-mo-" mozaika, a ime roda Reovirus je skraćenica riječi "dišni, respiratorni"- "R", "e" od enteric"- trbušni tifus i "o" od "orphan"- siroče". Ukoliko postoji više od jednog naziva za isti takson, odluka o tome koje ime će biti prihvaćeno, vrši se na temelju zakona ili, ako je potrebno, na osnovu prihvatljivosti za većinu virologa. Ako nije predloženo odgovarajuće ime za takson, takson može biti odobren, a ime ostaje neriješeno sve dok ICTV ne predloži i prihvati prijedlog međunarodnog imena. Postoje i tzv. „privremena imena“, na primjer, vrsta virusa „koja izaziva hlorotične pjege na listovima soje“ nalazi se u još uvijek „neimenovanom“ rodu familije Caulimoviridae. Dok rod ne dobije ime, ova vrsta virusa ostaće označena kao "vrsta koja izaziva hlorotične pjege na listovima soje".
13.4. Pravila o vrstama
Vrsta virusa je definirana kao „politetička grupa virusa koja ima sposobnost replikacije loze i zauzima posebnu ekološku nišu“. Ukoliko je Pododbor ICTV nesiguran o taksonomskom statusu nove vrste ili o dodjeli nove vrste nekom rodu, nova vrsta se navodi kao provizorna (uslovna) vrsta u odgovarajućem rodu ili familiji. Imena „uslovnih“ vrsta kao svojti, ne smiju se duplicirati sa već odobrenim imenima i moraju biti izabrana tako da nisu blisko slična imenima koja su trenutno u upotrebi, imenima koja su bila u upotrebi u nedavnoj prošlosti ili imenima vrsta koje su konačne. Ime vrste se može sastojati od nekoliko riječi, ali se mora razlikovati od imena drugih taksona. Imena vrsta ne smiju se sastojati samo od imena domaćina i riječi "virus". Imena vrsta obično obuhvataju više od jedne riječi. Na primjer, imena biljnih virusa obično su konstruirana kao domaćin + simptom + "virus" (npr. virus nekroze duhana), dok, nasuprot tome, virusi u familiji Bunyaviridae obično se imenuju po lokaciji na kojoj je virus pronađen + "virus" (npr. Bunyamwera virus). Ime vrste mora osigurati odgovarajuću i jasnu identifikaciju vrste. Imena vrsta moraju biti osobita i ne smiju biti u obliku koji se lako može zamijeniti s imenima drugih taksona. Brojevi, slova ili njihova kombinacija, mogu se koristiti prilikom imenovanja vrsta kao epitet vrsta, ako se takvi brojevi i slova već naširoko koriste. Postojeća imena vrsta kao što su goveđi adenovirus A, goveđi adenovirus B itd., te i ljudski herpesvirus 1, ljudski herpes virus 2, ljudski herpes virus 3 itd. opravdavaju upotrebu slijedećih brojeva u nizu. Imena kao što su "22" ili "A7" nisu prihvatljiva.
13.5. Pravila o rodu
Rod je grupa vrsta koje dijele neke zajedničke karakteristike. Ime roda označava jednu riječ koja završava na ... virus. Odobrenje novog roda mora biti popraćeno odobrenjem tipa vrste.
13.6. Pravila o podfamiliji
Podfamilije čine grupu rodova koji dijele neke zajedničke karakteristike. Takson se koristiti samo u slučaju kada je potrebno riješiti složene hijerarhijske probleme. Ime podfamilije čini jedna riječ koja završava na ... virinae.
13.7. Pravila o familiji
Familija predstavlja grupu rodova (koji jesu ili nisu organizirani u podfamilije) koji dijele neke zajedničke karakteristike. Ime familije čini jedna riječ koja završava na ... viridae.
13.8. Pravila o redu
Red čini grupa familija koje dijele neke zajedničke karakteristike. Ime reda sastoji se od jedne riječi koja završava na ... virales.
13.9. Viroidi
Pravila
koja se bave klasifikacijom virusa, primjenjuju se i na klasifikaciju
viroida. Formalni nastavci za svojte viroida su riječ "viroid"
za vrste, sufiks "-viroid" za rodove, sufiks "viroinae"
za podfamilije (ako se ukaže potreba za ovim taksonom) i "-viroidae"
za familije.
Na primjer, viroid vretenastog gomolja svrstan je u rod Pospiviroid,
a pripada familiji Pospiviroidae.
13.10. Ostali podvirusni agensi
Sateliti i prioni nisu klasificirani kao virusi, ali se proizvoljno dodijeljuju klasifikaciji, kada naučnici iz pojedinih podrčja to smatraju korisnim.
13.11. Pravila za pravopis
U službenom korištenju taksonomije, prihvaćena imena redova, familija, podfamilija i rodova virusa pišu se kurzivom, a prva slova imena se kapitaliziraju. Imena vrsta se pišu kurzivom i prvo slovo prve riječi velikim slovom. Druge riječi se ne kapitaliziraju, osim ako su u pitanju lične imenice, odnosno dijelovi lične imenice. Primjer pravilno napisanog oblika je "Tobacco mosaic virus", a primjer nepravilnog oblika je „tobaco mosaic virus“. Obzirom da sutaksoni apstrakcije, kada se njihova imena formalno koriste, pišu se isključivo italicom i kapitaliziranim slovima. U formalnoj upotrebi, imenu taksona treba da prethodi izraz za taksonomsku jedinicu.
Na primjer, ispravan formalni opis različitih taksona je ... familija Herpesviridae ... rod Morbillivirus, .... rod Rhinovirus, .... vrsta Tobacco necrosis virus - virus nekroze duhana itd.
Sažetak 8. Izvještaja ICTV (2009.)
- U 7. Izvještaju lCTV (2000.) po prvi put je pojam vrste virusa opisan kao najniži takson u hijerarhiji virusnih taksona. Kao što je definirano u njemu, " virusi predstavljaju politetičku grupu virusa koja ima sposobnost replikacije loze i zauzima posebnu ekološku nišu“."Politetička grupa" predstavlja grupu čiji članovi imaju nekoliko zajedničkih svojstava, iako svi članovi te grupe ne moraju dijeliti određena zajednička svojstva. Drugim riječima, članovi vrsta virusa su definirani konsenzusom zajedničkih svojstava grupe. Vrste virusa razlikuju od viših taksona virusa, koji predstavljaju "univerzalne" grupe, i kao takve imaju definirane osobine koje su potrebne za članstvo. Virusi su stvarni fizički subjekti koji su nastali kao proizvodi biološke evolucije i genetike, dok su vrste virusa i viši taksoni apstraktni pojmovi nastali kao proizvodi racionalne misli i logike. Relacija virus/vrsta time predstavlja liniju sučeljavanja između biologije i logike.
- Virusi (uključujući izolate virusa, sojeve, varijante, vrste, podvrste, serotipove, itd.) trebaju, kad god je to moguće, biti dodijeljeni odgovarajućoj vrsti virusa. Obzirom da je veliki broj virusa još uvijek neadekvatno okarakteriziran, mnogi virusi su i dalje neraspoređeni.
- Sve vrste virusa moraju biti reprenzentirane sa najmanje jednim izoliratom virusa.
- Gotovo sve vrste virusa su članovi priznatih rodova. Nekoliko vrsta ostalo je neraspoređeno u svojim familijama, iako su jasno identificirani kao nova vrsta.
- Neki rodovi su članovi priznatih podfamilija.
- Sve podfamilije i većina rodova su članovi priznatih familija. Neki rodovi još uvijek nisu dodijeljeni familijama, u budućnosti oni se mogu ili pridružiti postojećim familijama ili će predstavljati novu familiju sa drugim neraspoređenim rodovima.
- Neke familije su članovi slijedećih priznatih redova: Caudovirales, Nidovirales i Mononegavirales.
- Hijerarhija priznatih taksona virusa je:(red), familija, (podfamilija), rod i vrsta
- Samo navedeni taksoni su priznati od strane ICTV. Ostale grupacije u posebnim okolnostima mogu biti korisne, ali nemaju priznato taksonomsko značenje. Slično tome je i pojam "kvazi-vrste" koja nema priznato taksonomsko značenje, iako predstavlja važan koncept u taksonomiji virusa.
- Stvaranje ili eliminacija, (re) imenovanje i (re) dodjela vrste, roda, (pod) familije ili reda virusa su taksonomski akti za koje je potrebno temeljno proučavanje i javna rasprava, što dovodi do službenog odobrenja punopravnog članstva ICTV. Nasuprot tome, imenovanje izolata virusa i njihova dodjela već postojećim vrstama ne smatra se taksonomskim aktom i stoga ne zahtijeva formalno ICTV odobrenje. Umjesto toga, njihovo imenovanje vrši se objavljivanjem rada u indeksiranim časopisima iz oblasti virologije, u kojem se opisuje dotični izolat. Ovakav način imenovanja odnosi se i na opis satelita virusa, viroida, agensa humanih spongiformnih encefalopatija (priona) i nekoliko vrsta virusa životinja i gljiva.
- Broj priznatih svojti za svako taksonomsko izdanje:
Godina |
Redovi Orders |
Familije Families |
Podfamilije Subfamilies |
Rodovi Genera |
Vrsta Species |
2009. |
6 |
87 |
19 |
348 |
2290 |
2008. |
5 |
82 |
11 |
307 |
2083 |
8. Izvještaj |
3 |
73 |
11 |
289 |
1903 |
- Pojava određivanja nukleotidne sekvence je izazvala revolucionarne promjene u biologiji i uvelike racionalizirala taksonomiju, uključujući i viruse. Univerzalna taksonomija virusa pruža klasifikacijske sheme koje su podržane na osnovu provjerenih podataka i koncenzusa stručnjaka. To je neophodan okvir za daljnje istraživanje trenutno priznatih vrsta virusa i za identifikaciju i karakterizaciju novih virusa u nastajanju, bilo da su nastale kao posljedica prirodnog, slučajnog ili namjernog širenja.
LITERATURA
- Büchen-Osmond, C. (2003) Taxonomy and Classification of Viruses. Virology.
- Butler, P. Jonathan G., & Klug, A. (1978) The assembly of a virus. Scientific American239(5): 62–69, (Offprint No. 1412).
- Diener, T. O. (1981) Viroids. Scientific American244(1): 66–73, (Offprint No. 1488).
- Đermić, E. (2009) Biljna virologija i fitopatogene bakterije – skripta. Agronomski fakultet Sveučilišta u Zagrebu.
- Enquist, F., Skalka, R. (2004). Principles of Virology, second edition. ASM Press, Washington, DC.
- Fauquet, C., Stanley, J. (2003) Geminivirus classification and nomenclature: progress and problems. Association of Applied Biologists, 142:165-189.
- Fraenkel-Conrat, H. (1994) Rebuilding a virus. Scientific American194(6): 42–47, June (Offprint No. 9).
- Levy, J., A., Fraenkel-Conrat H., & Owens, R. (1994) Virology.