
OSTALI SEMINARSKI RADOVI
IZ BIOLOGIJE : |
|||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||

ĆELIJSKE ORGANELE I GOLDŽIJEV APARAT
Ćelijske organele
Ćelijske organele su djelovi, to jest komponente citoplazme preko kojih
ćelija vrši većinu svojih aktivnosti a koje imaju sopstvenu veličinu,
oblik, funkciju i strukturu. Djelimo ih u dvije grupe:
a) membranske - specijalizovane komponente koje su sopstvenom
membranom odvojene od citosola. U ovu grupu organela spadaju: Endoplazmin
retikulum, Goldžijev aparat, lizozomi, peroksizomi i mitohondrije.
b) nemembranske - nisu odvojene od citosola s obzirom
da ne posjeduju sopstvenu memranu. U nemembranske organele spadaju centrioli
i ribozomi.

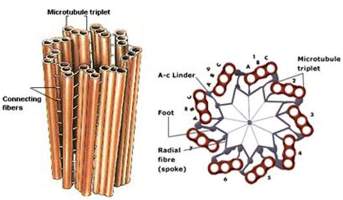
Centrioli su prečnika 0.2 µm, dužine oko 0.4 – 0.5µm. Cilindrične su strukture sa elekrono-gustim zidom i elektrono-svjetlom centralnom šupljinom. Zid centriola izgrađen je od 9 tripleta djelimično fuzionisanih mikrotubula - A, B i C (izgrađenih od β – tubulina) koji su poređani u krug. Sa prečnikom centriola mikrotubuli zaklapaju ugao od 50°. Znaćaj centriola je u tome što formiraju deobeno vreteno, mikrotubule citoplazme i aksoneme kod cilija i flagela. Ova organela se ne dijeli vec se replikacija ( koja počinje u S fazi ćelijskog ciklusa) odvija tako što uz svaki centriol izrasta novi pa od jednog para centriola nastaju dva para koji migriraju ka suprotnim polovima obrazujući deobeno vreteno. Bazalno tijelo se naziva specijalizovani centriol koji formira aksonemu treplji.
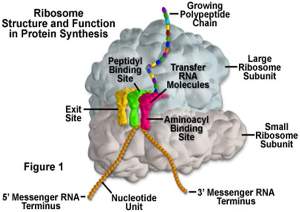
Ribozomi su dimenzija 20x30 nm, prisutni u svim ćelijama osim u eritrocitima, sitne, elektrono-guste organele od kojih oko 60% saržaja čine ribozimske RNK a ostatak 70 različitih strukturnih proteina i enzima potrebnih za sintezu proteina. Građeni su od manje subjedinice koja sadrži jedan molekul rRNK i 33 različita proteina i ima koeficijent sedimentacije 40, i veće subjedinice koja sadrži 3 molekula rRNK i 40 proteina i ima koeficijent sedimenatcije 60. Između subjedinica pri spajanju ostaje pukotina kroz koju klizi lanac informacione RNK. Tinktorijalnlo svosjtvo ribozoma zapaženo je još u 19.vijeku a nastaje kao posljedica bazofilije ribozoma (bazofilni regioni nazvani su: Nissl-ova supstanca u neuronima ili bazofilna tjelašca u čelijama angažovanim za sintezu proteina i ergastoplazma u žljezdanim ćelijama) koju prouzrokuje rRNK čiji molekuli sadrže mnoštvo fosfatnih grupa koji dejstvuju kao polianjoni. Monoribozomi su pojedinačni ribozomi a postoje i polozomi-poliribozomi koji su nanizani duž molekula iRNK i čine niz od 3-15 ribozoma raspoređenih na rastojanju od 15nm. Polizomi mogu biti slobodni u citosolu ili vezani za membranu ER-a, ali između njih nema strukturnih razlika. Na slobodnim polizomima se vrši sinteza proteina koji ostaju u ćeliji-a)citosolni proteini(hemoglobin, kontraktilni proteini, keratin); b)periferni memranski proteini i c)većina mitohondrijskih enzima. Na vezanim polizomima nastaju: a)proteini koje ćelija izlučuje, b)proteini koji se deponuju u ćeliji, c)integralni membranski proteini, ubacuju se u lumen ER-a i preko transportnih vezikula prenose do Goldžijevog aparata u kome se vrši modifikovanje.
Endoplazmin retikulum je izgrađen od unutarćelijskih šupljina raspoređenih u lavirint, a odvojenih od citosola sopstvenom mebranom, koje mogu imati spoljšten(cisterne), ovalan(vezikule) ili cjevast(tubuli) izgled, ili biti u kombinaciji pomenutih oblika, a anostomozovane su međusobno tako da grade mrežu sa jedinstvenim lumenom i otuda i naziv (lat. rele-mreža).
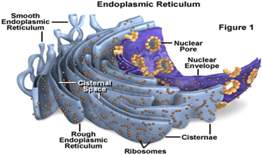
Postoje dva oblika ER-a: granulisani i glatki ER.
a) Granulisani (hrapavi) ER sastoji se od spoljštenih paralelno raspoređenih
cisterni, ima sposobnost sinteze proteina jer sadrži membranske proteine
riboforin I i riboforin II zbog čega može da vezuje ribozome i prihvata
novosintetisane proteine. Sadrže je sve ćelije osim eritrocita,
a zbog sadržaja ribozoma uzrokuje bazofiliju citoplazme. Na ribozomima
se sintetišu proteini namjenjeni za sekreciju ili same potrebe ćelije
(enzimi lizozoma,proteinske komponente plazmaleme, jedrovog omotača i
membrane ćelijskog omotača). Informaciona RNK odgovorna za sintezu proteina(koja
počinje na slobodnim polizomima a ubacuju se u cisternu grER-a), početnog
kodona sadrži kratak segment lanca koji kodira niz od 20-25 aminokiselina
koji se nazivaju signalna sekvenca (signalni peptid). Čestica za raspoznavanje
(signal recognition particle-SRP) je nukleoproteinski kompleks koji se
vezuje za signalnu frekvencu i sprečava dalju elongaciju polipeptidnog
lanca, vezuje se za odgovarajući receptor na membrani grER-a i omogućava
pričvrćivanje veće subjedinice za receptor riboforin nakon čega difunduje
u citosol a translacija se nastavlja. Enzim signalna pepdaza djeluje na
polipeptidni lanac u lumenu grER-a tako što cijepa signalnu sekvencu.
Novosintetisani protein mogu se modifikovati (hodralacija, sulfatacija,
glikozilacija, fosfatacija polipetidnog lanca) I deponovati ili transportovati
ka Goldži aparatu.
b) Glatki ER-sadrži mrežu bogato anastomozovanih cjevastih šupljina, širine
20-60nm koji nije u vezi sa ribozomima. Posjeduje membranski sistem koji
je povezan sa membranama gr ER-a i cisternama Goldži aparata u kojem se
nalaze enzimi koji su odgovorni za sintezu lipida i unutarćelijski transport,
što i jesu glavne uloge glatkog ER-a, ali ne i jedine. Oksidacija, metilacija
i konjugacija pojedinih hormona, metabolita, ljekova ili štetnih materija
odvijaju se u hapatocitama a u ovim procesima učstvuje i glER. Učestvuje
i u metabolizmu glikogena i zbog toga je glER najzastupljeniji u ćelijama
kore nadbubrega i polnih žlijezda i jetre. U skeletnim i srčanim mišićima
glER se označava kao sarkoplazmin retikulum jer se u njegovoj memrani
nalazi pumpa Ca(2+) jona koji su neophodni za kontrakcije mišićnih ćelija.
Lizozomi su zaduženi za intracelularnu digestiju tj. razlaganje ćelije, njenih djelova, bakterija, virusa, sopstvenih istrošenih komponenti. Heterogene su u pogledu veličine, oblika i elektrone gustine, prisutni u svim ćelijama osim u eritrocitima, djele se na primarne sekundarne i tercijarne.
Primarni lizozomi(vezikule sferičnog ili ovalnog oblika, prečnika 5-50nm)
su uključeni u intracelularnu digestiju, skladištenje i transportovanje
hidrolaznih enzima. Sadrže preko 40 tipova kiselih hidrolaza (glikozidaze,
nukleaze, proteaze, fostfataze itd.) koji mogu razložiti većinu biomolekula.
Optimalna pH je 5. Membrana sprečava da enzimi iscure u citosol i razgrade
citoplazmu a otporna je na dejstvo proteolitičkih enzima jer je visoko
glikozilovana. Sekundarni lizozomi(nepravilan oblik, dimenzije 0.2-2µm,)
nastaju spajanjem primarnih lizozoma sa fagozomima – fagolizozomi, sa
autofagozomima – autofagolizozomi, i kasnim endozomima. Vrše degradaciju
supstanci unijetih u ćeliju ili sopstvenih dotrajalih organela, čiji se
produkti izbaciju u citosol a potom egzocitozom u ekstracelularni prostor.
Tercijarni lizozomi ili rezidualna tijela jesu nesvareni ostaci zadržani
u ćeliji ćcijom fuzijom nastaje pigment starenja – lipofucin.
Multivezikulna tjelašca se smatraju varijantom sekundarnih lizozoma u
kojima se razgađuje veći broj endocitoznih vezikula, ili membranski sistem
za transponovanje materijala do kasnih endozoma, a predstavljaju vakuole
nepravilnog oblika u čijem matriksu se uočava veći broj prividno praznih
vezikula. Autoliza jeste patološko stanje kada dođe do prskanja membrane
i oslabađanja kiselih hidrolaza u citosol i samim tim liziranja citoplazme
sopstvene ćelije. Ova pojava samouništenja ili apoptoze jetse fiziološka
reakcija organizma kojim se obezbjeđuje formiranje tkiva i organa u toku
embriogenze i omogućava održavanje homeostaze u postnatalnom periodu.
Peroksizomi (sferične ili ovalne strukture, prečnika 0.2-1µm) . S obzirom da na elektronskom mikroskopu pokazuju slične karakteristike kao i primarni lizozomi, razlike(na osnovu morfoloških kriterijuma) je moguće prikazati histohemijskim tehnikama jer peroksizomi sadrže drugačiji set enzima.

Sadrže enzime koji se sintetišu u citosolu na slobodnim ribozomima a zatim transportuju kroz membranu peroksizoma i djele se prostom diobom (fisijom). Najznačajniji su peroksidaza, oksidaza, D-aminokiselina, katalaza. Peroksizomi su najzastupljeniji u hepatocitama i nefrocitama zbog uloge u detoksikaciji i metabolizmu lipida. Pri katabolizmu masnih kiselina nastaje vodonik – peroksid (H2O2), koji koriste hepatocite i nefrocite pri detoksikaciji(oksidacija toksičnih materija), a fagociti za uništavanje mikroorganizama. U višoj koncentraciji može biti štetan zbog čega peroksizomi sadrže enzime- katalaze koji ga razlažu na vodu i kiseonik (2H2O2=2H2O + O2).
Mitohondrije (najčešće štapićast izgled, širine 0.4-0.8µm a dužine 4-8µm, prisutne u svim ćelijama osim u eritrocitima i keratinocitama). Promjenjivog su oblika i dimenzija a uloga im je u tome što generišu energiju koja je neophodna za biosinteteske i motorne aktivnosti ćelije pa im je broj uslovljen stepenom aktivnosti ćelije (brojne su u srčanom mišiću, hepatocitama(može imati 2000 mitohondrija), nefrocitama, ivičnim ćelijama želudačnih žlijezda, a najbrojnije su u oksifilnim ćelijama paratiroidne žlijezde). U građi mitohondrija razlikuju se spoljašnja membrana, unutrašnja membrana, i matriks(unutrašnja komora).
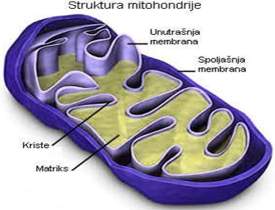
Spoljašnja membrana predstavlja relativno propusnu opnu širine 6-7 nm,
a funkcija joj je da odvaja mitohondriju od citosola. Proteini porini
formiraju vodene kanalice kroz membranu, kroz koje difunduju joni i hidrosolubilni
molekuli (mase do 10kDa) i stvaraju intermembranski prostor sličan citosolu.
Osim proteina porina membrana sadrži i enzime koji učestvuju u konverziji
masnih kiselina koje se metabolišu u matriksu organele. Unutrašnja membrana
je tanja od spoljašnje- 5-6 nm, ali je neporpusnija što je posljedica
prisustva velike količine kardiolipina – fosfolipidnog molekula koji posjeduje
glavu i četiri repa(četiri masne kiseline). Kontaktna mjesta su regioni
u kojima su dvije mitohondrijske membrane spojene i pomoću prisutnih proteina
u membrani ostvaruje se razmjena metabolita kao i transport elektrona
(pomoću proteinskih kompleksa). Lanac za transport elektrona čini niz
proteinskih kompleksa koji primaju i otpuštaju elektrone i time se redukuju
i oksidišu, a ovaj lanac čine flavoprotein, proteini koji sadrža gvožđe-sulfid,
ubikvinon i citohromi B, C, C1, A, A3. Pri prolasku elektrona kroz respiratorni
lanac-stvara se energija koju komponente koriste za transport elektrona
iz mitohondrijskog matriksa u intermembranski prostor i izmedju ova dva
dijela se stvara visok elektrohemijski gradijent.
Elementarna tjelašca(oksizomi, ATP-sintetaze) su u obliku sijalice i kroz
njih difunduju H+ joni i protok ovih jona oslobađa energiju za sintezu
ATP-a. Intermembranski prostor predstavlja prostor izmedju spoljašnje
i unutrašnje memrane , širok je 10-20 nm, a u kristama(nabori na unutrašnjoj
membrani, duplikature koje sadrže enzime i molekule uključene u proces
fosforilacije) se širi i zavlači u duplikature unutrašnje membrane. Mitohondrijski
matriks (unutrašnja komora) – prostor koji je ograničen unutrašnjom membranom,
sadrži enzime, mitohodrijalnu DNK i RNK, ribozome i matriksne granule.
Ćelijske inkluzije (produkti ćelijskog metabolizma) mogu biti u obliku kapljica, granula ili kristala, i u tom obliku se deponuju u citoplazmi. Neke inkluzije kao npr. pigmentne granule mogu biti obavijene membranom, dok je većina uronjena u citoplazmu. Najznačajnije inkluzije su: -glikogen – polisaharid koji nastaje u glER-u od glukoze. Kod ćelija sa izraženom metaboličkom aktivnošću veoma je izražen jer je bogat energijom. Na svjetlosnom kikroskopu vidi se tek nakon bojenja. -masne kapljice- depoziti triglicerida kao izvora energije, i holesterola kao prekusora steroidnih hormona. Adipociti su velike kapi dok mnoštvo sitnih kapljica daje saćast izgled ćeliji. -pigmenti su depoziti obojenih supstanci i mogu biti intracelularni ekstracelularni. Egzogeni pigmenti se u tijelo unose iz spoljašnje sredine i u tkivima se ponašaju kao strana tijela ali nemaju uticaja na osnovne metaboličke procese. Kao na primjer ugljena prašina koja je prisutna u makrofagima pluća odakle se prenosi do limfnih čvorova respiratornog sistema. Endrogeni pigmenti su specifični produkti neke ćelije. Mnogi od njih su vezi sa ćelijom u kojoj su deponovani dok neki predstavljaju rezultat starenja. Ovdje spadaju: melanin, hemoglobin, mioglobin, fotohemijske supstance, rodopsin, jodopsin, citohromi, lipogeni pogmenti kao lipofuscin, lipohrom, luteini.
GOLDŽIJEV APARAT
Drugačije se naziva Goldžijev kompleks , spada u membranske organele.
Otkriven je u nervnim ćelijama još u 19.vijeku od strane italijanskog
biologa K.Goldži-ja (Camillo Golgi). Uključen je u sintezu ugljenih hidrata
i lipida kao i modifikovanje, razvrstavanje i usmjeravanje protein iz
ER-a, prisutan je u svim ćelijama osim u eritrocitima i orožalim ćelijama
epidermisa. Na svjetlosnom mikroskopu , na preparatima bojenim hematoksilin
– eozionom, uočava se kao mreža smještena u blizini jedra. Goldžijev aparat
je građen od 3-10 spljoštenih, paralelno raspoređenih diskoidnih vrećica
ili cisterni, što se može zapaziti na elektronskom mikroskopu. Cisterne
su razdvojene prostorom od 5-30nm tako da ne komuniciraju međusobno. Što
se tiče građe cisterni, središnji dio im je uzan dok su krajevi prošireni.
One ispoljavaju morfološku i funkcionalnu polarizovanost. Razlikuju se
konveksna od konkavne strane organele. Konveksna strana je okrenuta prema
jedru ili ER-u i ona je formirajuća ili cis strana, dok je konkavna strana
okrenuta prema čelijskoj membrani i ona se označava kao maturirajuća ili
trans strana. Mrežast izgled je posljedica usmjereno fenestrovanih cisterni
u tolikoj mjeri da dobijaju oblik anastomozovanih tubula.
Na ispupčenoj strani nalaze se cis Goldžijeve cisterne dok se na udubljenoj
strani nalaze trans Goldžijeve cisterne a između njih se nalaze intermedijerne
cisterne.
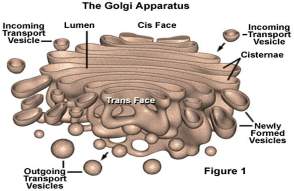
Na elektronomikrografijama Goldžijev aparat često se ne razlikuje od
gl ER-a, jer su memrane cisterni glatke, mada se uz cisterne zapaćaju
vezikule i vakuole razlićite velićine i elektronske gustine. Izgled Goldžijevog
aparata varira u odnosu na aktivnosti same ćelije , tako da se na eletronomikrografijama
ne uoćava ćesto klasićna forma organele. Goldžijevo polje ćine nekoliko
povezanih Goldžijevih aparata u jednoj čeliji, a u jednom Goldžijevom
polju moće biti čak 100 cisterni. Preko transportnih vezikula proteini
i lipidi sintetisani u ER-u prenose se do cis Goldžijeve mreže (konveksna
strana nalazi se u blizini ER-a), i tansportuju u pravcu trans Goldžijeve
mreže. U cisternama proteini i lipidi se modifikuju- glikoziluju čime
nastaju glikoproteini, glikolipidi i proteoglikani. Ovako modifikovani
ili novosintetisani proteini se pakuju u sekretne vezikule, vakuole ili
primarne lizozome i otpremaju ka konačnim odredištima. Sekretne vezikule
imaju glatku površinu a način na koji one transportuju materije naziva
se konstitutivna sekrecija – bez zadrćavanja u citoplazmi one se kreću
ka plazmalemi i oslobađaju sadržaj van ćelije. Slično fibroblasti izlučuju
glikoproteine ekstracelularnog matriksa, peharaste ćelije sluz itd. Sekretne(kondenzujuće)
vakuole se na elektronskom mikroskopu uočavaju kao čekinjaste zbog toga
šsto su obložene klatrinskim omotačem. Mehanizam sekrecije kod ovih vakuola
naziva se regulisana sekrecija. Funkcioniše na načcin što nakon odvajnja
od trans mreže Goldžijevog aprata one se zadržavaju u citosolu pri čemu
se njihov sadržaj zgušnjava i koncentriše i do 100 puta. Sekretne granule
se nazivaju vakuole sa koncentrovanim sadržajem i oslobađaju svoj sadržaj
jedino usljed adekvatne stimulacije. Ovaj vid sekrecije svojstven je ćelijama
koje luče digestivne enzime, hormone ili neurotransmitere. Materije se
oslobađaju egzocitozom, a Goldžijev aparat indirektno učestvuje u obnavljanju
plazmaleme tako što se membrane vezikula i granula stapaju sa njom i postaju
dio nje. U cis Goldžijevim citernama za solubilne lizozome vezuju se grupe
manozo-6-fosfata. U membrani trans Goldžijeve mreže prisutan je manozo-6-fosfat
(M-6-F receptor)i vezuje M-6-F. Ovaj proces naziva se fosforilacija manoznih
ostataka i on prethodi razvrstavanju lizozomskih enzima (kiselih hidrolaza)
. Primarni lizozomi nastaju pupljenjem klatrinskih vezikula ispunjenim
hidrolazama, a pupljenju pomaže klatrinom obložena spoljašnja strana trans
Goldžijeve mreže. Reciklaža receptora jeste proces vraćanja djelova lizozoma
koji sadrže M-6-F receptore u trans Goldžijevu mrežu, posle stapanja sa
vezikulama čiji se sadržaj razgrađuje. Primarni lizozomi gube klatrinski
omotač po odvajanju od Goldžijevog aparata. Kod ćelija koje sadrže veliku
količinu proteina i glikoproteina kao što su ćelije pljuvačnih žljezda,
pankreasa, plazmociti, osteoblasti, kao i ćelije koje često obnavljaju
plazmalemu - neuroni , Goldžijev aparat je prisutan u velikoj količini.
Perinukleusno rasvetljenje ili kako se tradicionalno naziva, perinkleusni
halo jeste kako se Goldžijev aparat uočava na svjetlosnom mikroskopu s
obzirom da se ne boji hematoksilin-eozinom.
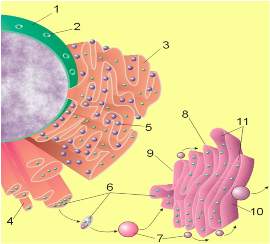 1.
Nuklearna memrana
1.
Nuklearna memrana
2. Nuklearne pore
3. Granularni endoplazmin retikulum (grER)
4. Glatki endoplazmin retikulum (glER)
5. Ribozomi na grER-u
6. Makromolekuli
7. Transportne vezikule
8. Goldžijev aparat
9. Cis strana Goldži aparata
10. Trans strana Goldži aparata
11.Cisterne Goldži aparata
Literatura:
• Ćelija i tkiva , dr Zlatibor Anđelković
• Babić, M. i saradnici. Medicinski leksikon. Zagreb: 1999.
• Grujčić-Milović, S. i Tomašić, Lj. Atlas hematologije i citologije. Zagreb: 2001.
• Grozdanović-Radovanović, J. Citologija. Beograd: 2004.
• Guyton, A. C. i Hall, J. E. Medicinska fiziologija. SAD: 1992.
• Melita, Š. Citologija – priručnik za nastavnike. Sarajevo: 1976.
• Sobota, J. Histološki atlas–citologija, histologija, mikroskopska anatomija. Zagreb: 2004.
• Hristić, M. i Potparević, B., Praktikum iz Biologije sa Humanom genetikom.
preuzmi
seminarski rad u wordu » » »
